Habitat/Ökologie (Meinunger & Schröder 2007) Eine innerhalb der Familie der Lophoziaceae systematisch isoliert stehende, wenig veränderliche und daher leicht bestimmbare Art. Kalkmeidende Art an meist halbschattigen, luftfeuchten Stellen in lockeren Rasen oder Decken, auch einzeln zwischen anderen Moosen. Auf feuchtem, humosem, saurem Waldboden, an Wegböschungen, in beerstrauchreichen Nadelwäldern, an Felsen und in Blockhalden, in den Alpen besonders auf Latschenhumus. Begleiter sind: Rhytidiadelphus loreus, Plagiothecium undulatum, Dicranum scoparium, D. fuscescens, Mylia taylorii, Barbilophozia attenuata, Bazzania trilobata, Dicranodontium denudatum, Tetraphis pellucida, Lepidozia reptans, Lophozia ventricosa, Campylopus flexuosus. Angaben zu Soziologie und Ökologie bei Matzke (1990); Marstaller (1991a); Mohr (2002); F. Müller (2004); Nebel & Philippi (2005). Lophozia ventricosa agg.: Taxonomischnomenklatorische Vorbemerkungen: Die Gliederung der Lophozia ventricosa-Gruppe ist bis in die neueste Zeit unklar und verworren. Bei vielen Proben kommt man, je nachdem welche Literatur man benutzt, zu unterschiedlichen Bestimmungsergebnissen. Wir haben uns jahrelang mit dieser Gruppe befasst und dabei tausende Proben untersucht, wobei wir dazu übergegangen sind, die Proben nach klaren Kriterien zu sortieren, ohne uns zunächst auf bestimmte Namen zu fixieren. Die Arbeiten erfolgten in enger Abstimmung und Diskussion mit H. Köckinger, der die Verhältnisse in den österreichischen Alpen untersucht und dem auch an dieser Stelle nochmals gedankt sei. Ferner danken wir R. Grolle und H.-J. Zündorf (Jena), die uns die Möglichkeit gaben, Typusmaterial von Lophozia longiflora, Lophozia guttulata sowie den Neotypus Lophozia ventricosa Grolle & Long (2000) einzusehen und zu untersuchen. Wir sind zu dem Ergebnis gekommen, dass sich diese Gruppe in unserem Gebiet in die vier Arten Lophozia ventricosa, L. wenzelii, L longiflora und L. guttulata zerlegen lässt und dass sich jede einigermaßen gut entwickelte Probe auch ohne Kenntnis der Ölkörper eindeutig bestimmen lässt. Lophozia ventricosa agg. umfasst mittelgroße, diözische Pflanzen mit hyalinen, ungefärbten Brutkörpern. Nur bei stark rot gefärbten Pflanzen in offenen, meist alpinen Lagen können die Zellwände der Gemmen bisweilen leicht rosarot eingefärbt sein. Die habituell ähnliche und ebenfalls sehr formenreiche Lophozia sudetica unterscheidet sich durch (reif) braunrote Brutkörper, Übergangsformen zwischen beiden, von denen in älterer Literatur öfter die Rede ist, gibt es nicht. Anerkanntes Typusmaterial zu Jungermannia ventricosa Dicks. scheint nicht mehr vorhanden zu sein oder wurde zumindest von späteren Taxonomen deshalb nicht in Betracht gezogen, weil über die Beschaffenheit der Ölkörper keine Informationen vorliegen: Schuster (1969); Grolle & Long (2000). Eine erste zusammenfassende Darstellung für unser Gebiet findet sich bei K. Müller (1905–1916). Hier werden innerhalb dieser Gruppe Lophozia wenzelii, L. confertifolia, L. guttulata, L. longiflora und L. ventricosa als Arten unterschieden. Als trennende Merkmale werden vor allem Blattform, Größe der Pflanzen und unterschiedliche Standorte verwendet. Liest man die ausführlichen Kommentare zu diesen Arten, so geht daraus hervor, dass bei vielen Proben eine klare Zuordnung nicht möglich ist. Diese Unsicherheiten setzen sich in der nachfolgenden, insbesondere auch floristischen Literatur, für unser Gebiet fort. Einige Jahrzehnte später hat sich H. Buch (1933) eingehender mit dieser Gruppe befasst und eine weitere neue Art, Lophozia silvicola Buch (1933a) beschrieben. Diese ist gekennzeichnet durch bikonzentrische Ölkörper und lässt sich scheinbar leicht und bequem bestimmen. Leider stellte sich bald heraus, dass diese zunächst bestechende Methode mit schweren Nachteilen behaftet ist, zum einen ist getrocknetes Herbarmaterial damit auch weiterhin nicht bestimmbar, zum anderen gibt es Pflanzen, in denen neben bikonzentrischen gleichzeitig auch homogene Ölkörper auftreten sowie „Zerfallsstufen“ dazwischen. Untersucht man eine hinreichend große Anzahl von Proben, kann man alle Übergangsformen zwischen echter Lophozia silvicola bis hin zu normaler L. ventricosa finden. Erste Zweifel in dieser Hinsicht wurden bereits bei Schade (1936) angemeldet, und neben anderen Bryologen haben auch wir solche Beobachtungen gemacht. Schuster (1969) hat ein weiteres Konzept dieser Gruppe entwickelt. Darin wird L. silvicola wieder als Varietät zu L. ventricosa gestellt. Als Unterscheidungskriterien werden weiterhin hauptsächlich Blattmerkmale herangezogen. Dieses Konzept wird dann von den meisten Nachfolgern, jeweils mit einigen Variationen, übernommen, so von Paton (1999), Schumacker & Váña (2000), Damsholt (2002) und Grolle & Long (2000). Überblickt man diese Arbeiten, stellt man fest, dass sich zwar die Zahl der taxonomischen Einheiten auf unterschiedlichen Hierarchiestufen weiter erhöht hat, die Bestimmungsunsicherheiten aber weiter unverändert geblieben sind, wie dies etwa in den kritischen Anmerkungen bei Grolle & Long (2000) deutlich zum Ausdruck gebracht wird. Ursache für diese unbefriedigende Situation ist offenbar die große Variabilität der bisher verwendeten Merkmale, die sich stark überlappen und eine klare Unterscheidung auf Artniveau nicht gestatten. Ein Ausweg ist nur in Sicht, wenn man versucht, das Material mit weiteren, bisher nicht benutzten brauchbaren Unterscheidungskriterien neu zu gliedern. Ein erster Schritt in diese Richtung findet sich bereits bei K. Müller (1951–1958), der im Text darauf hinweist, dass sich Lophozia ventricosa und L. wenzelii durch Unterschiede in den Stängelquerschnitten klar trennen lassen, dieses Merkmal jedoch im Schlüssel nicht verwendet. Als Erster hat Saukel (1985) Unterschiede in den Stängelquerschnitten konsequent und mit überraschend gutem Erfolg bei der Neugliederung dieser Gruppe herangezogen. Leider ist diese wichtige Arbeit in der danach erschienenen, oben zusammengestellten Fachliteratur nirgends zitiert und fand keinerlei Beachtung, was nur als schwerwiegendes Versäumnis bezeichnet werden kann. Wir haben unser Material nach diesem Schlüssel bestimmt und konnten seine Ergebnisse weitgehend bestätigen: Meinunger (2001). In der Zwischenzeit haben wir dort noch unklare Fragen weiter verfolgt und hatten Gelegenheit, Typusmaterial zu einigen Arten zu untersuchen. Nach unseren derzeitigen Erkenntnissen lässt sich der Komplex nach den Stängelquerschnitten in zwei klar abgegrenzte Gruppen einteilen (vergl. Abb. 21): A) Stängelquerschnitt in der unteren Hälfte bis etwa zur Mitte mit meist 5–20 Lagen kleinzelliger Zellen, die deutlich von den darüber (außen) liegenden größeren Zellen abgegrenzt sind: K. Müller (1951–1958, Fig. 195a); Saukel (1985, Abb. 3a, 3b). B) Stängelquerschnitt meist nur mit 1–4 Lagen kleinzelliger Zellen über den Rindenzellen im unteren Teil, höchstens bis ins untere Drittel des Stängelquerschnittes reichend: K. Müller (l. c., Fig. 195c); Saukel (1985, Abb. 3c). Die Schnitte müssen im Mittelteil gut entwickelter, (möglichst) steriler Triebe gemacht werden. Sie müssen senkrecht zur Sprossachse erfolgen und hinreichend dünn sein. Meist sind die kleinzelligen Zellen verpilzt und dann farblich deutlich von den übrigen Zellen abgesetzt. Dieses in der Literatur oft bemühte Merkmal der Verpilzung ist zwar auffällig, aber nicht entscheidend. An zu jungen Sprossteilen sind auch die kleinzelligen Zellen oft noch unverpilzt und hyalin. Auch die größeren, darüber liegenden Zellen können teilweise verpilzt sein, bei L. wenzelii ist dies sogar regelmäßig der Fall. Wir haben mehrere hundert Proben geschnitten und keine Übergänge gefunden, diese Ergebnisse wurden inzwischen auch von anderen Bryologen bestätigt. Danach lassen sich nach folgendem Schema vier Arten klar abgrenzen.
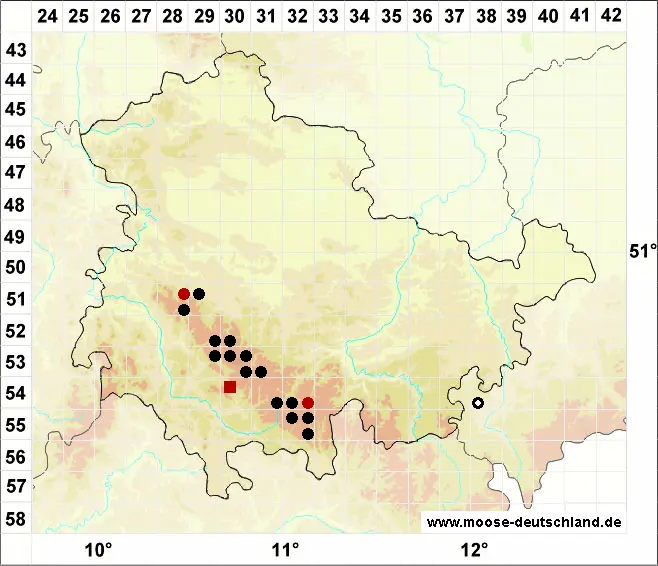
Verbreitung (Meinunger & Schröder 2007) Nur in den höheren Mittelgebirgen und in den Alpen, sehr selten in engen, luftfeuchten Tälern tiefer herabsteigend: NB, ST: Nur im Oberharz, vor allem im Brockengebiet: Loeske (1903); Philippi (1963); Marstaller (1991a). Von vielen neueren Bryologen bestätigt, u. a. (!!). (NW): Unklare Angabe: F. Koppe (1977), nach Schmidt & Heinrichs (1999) zu streichen. TH: Zerstreut in höheren Lagen des Thüringer Waldes: Meinunger (1992). SN: Selten im Westerzgebirge und im Vogtland: F. Müller (2004). RP: Nur an wenigen Stellen im Hunsrück: Matzke (1990); Caspari (2004). BW: Nur in hohen Lagen des Schwarzwaldes: Nebel & Philippi (2005). BY: Nordbayern: Vereinzelt im Fichtelgebirge und Steinwald: Hertel & Wurzel (2006 !!); in Hochlagen des Bayerischen Waldes vielfach, vereinzelt in engen Tälern zur Donau hin: Familler (1917); F. Koppe (1955); F. & K. Koppe (1931); Dürhammer (1997); Mohr (2002); M. Reimann (in litt. !!). Alpen: In Bergwäldern um die Waldgrenze ist die Art ziemlich verbreitet.
Bestand und Gefährdung (Meinunger & Schröder 2007) In den Alpen, im Schwarzwald und im Bayerischen Wald kann es zur Ausbildung größerer Bestände kommen, hier ist die Art derzeit ungefährdet. Im Harz, Thüringer Wald, Fichtelgebirge und Hunsrück sind die Bestände kleiner und weniger vital, hier muss die Art als gefährdet eingestuft werden: RL 3. Im Erzgebirge sind nur noch kleine, oft kümmerlich entwickelte Bestände vorhanden, nach F. Müller (2004) hier RL 1. Eine große Gefahr für die Art geht neuerdings von Waldkalkungsmaßnahmen aus, gut entwickelte Bestände von Anastrepta orcadensis müssen davon unbedingt verschont und die Bestände geschützt bleiben.




 [ Verbreitung in Deutschland ]
[ Verbreitung in Deutschland ]