Habitat/Ökologie (Meinunger & Schröder 2007) Ein variabler, im Gelände unauffälliger Artkomplex, den man sich erst durch umfangreiche Vergleiche mit ähnlichen Arten erarbeiten muss, um Bestimmungssicherheit zu bekommen. Zu empfehlen ist die Darstellung in Nyholm (1979). Die Pflanzen sind meist hell- bis gelbgrün und kräftiger als die Vertreter von Amglystegium serpens agg. Diagnostisch wichtig ist das verlängertprosenchymatische, aber lockere Zellnetz. Die Stängelblätter sind aus breit eiförmigem Grund schmal lanzettlich lang zugespitzt, sie laufen am Stängel nicht oder nur kurz und undeutlich herab. Dadurch unterscheidet sich die Art sicher von Amblystegium radicale mit deutlich herablaufenden Blättern und insgesamt schmaleren Zellen. Von ähnlichen Formen von Drepanocladus aduncus und Campylium polygamum unterscheidet sie sich durch die fehlende, vergrößerte hyaline oder gelbe Blattflügelzellgruppe (gut entwickelte Stammblätter untersuchen!). Die Art zerfällt in die zwei „Grundtypen“ Amblystegium kochii und A. trichopodium, die zueinander etwa im gleichen Verhältnis stehen wie A. serpens und A. juratzkanum, gute Abbildungen dieser Formen bei Roth (1905). Bei Amblystegium kochii sind die Blätter breit oval und ganzrandig, das Zellnetz überall locker und relativ breit, die wenig gebogene Blattrippe endet in oder wenig oberhalb der Blattmitte. Bei A. trichopodium sind die Blätter schmaler und spitzer, das Zellnetz ist am Grunde locker wie bei A. kochii, gegen die Spitze wird es besonders am Rande deutlich schmaler und Leptodictyum riparium ähnlich. Die Rippe reicht weit hinauf, ist in der oberen Blatthälfte deutlich verbogen, wodurch das Blatt asymmetrisch erscheint, und verliert sich in der Blattspitze. Besonders die Astblätter sind meist deutlich gezähnelt. Zu A. trichopodium gehört nach uns vorliegenden Belegen auch A. varium var. paludosum (Hansen) Moenkem., Moenkemeyer (1927). Bei A. varium ist das Blattzellnetz auch im oberen Blattteil immer kurz, die Rippe ist immer gut begrenzt, und alle Blätter sind ganzrandig. Bei Leptodictyum riparium ist das Zellnetz auch im unteren Teil des Blattes schmal, und die Rippe ist immer gerade. Amblystegium kochii und A. trichopodium sind in typischer Ausbildung gut zu unterscheiden, doch scheinen beide durch Zwischenformen verbunden zu sein, so dass wir sie hier nicht getrennt behandeln. An feuchten bis nassen, meist basen- und nährstoffreichen Standorten. Vor allem auf Feucht- und Nasswiesen, auch auf Salzwiesen, in Kies- und Tongruben, Steinbrüchen, im Verlandungsbereich stehender Gewässer, auf feuchter Moorerde, in Erlenbrüchen und an feuchten Gräben. Begleitarten sind: Drepanocladus aduncus, Brachythecium rutabulum, B. rivulare, Calliergonella cuspidata, Leptodictyum riparium, Bryum pseudotriquetrum, Climacium dendroides, Eurhynchium hians. Wegen großer Bestimmungsunsicherheiten sind die bisherigen Aussagen zur Soziologie kritisch zu betrachten. So treten etwa A. humile und A. radicale nur ausnahmsweise gemeinsam auf. Amblystegium radicale ist an humide, stärker saure Standorte gebunden, während A. humile deutlich wärmeliebender ist und basenreichere, bisweilen recht trockene Standorte bevorzugt.
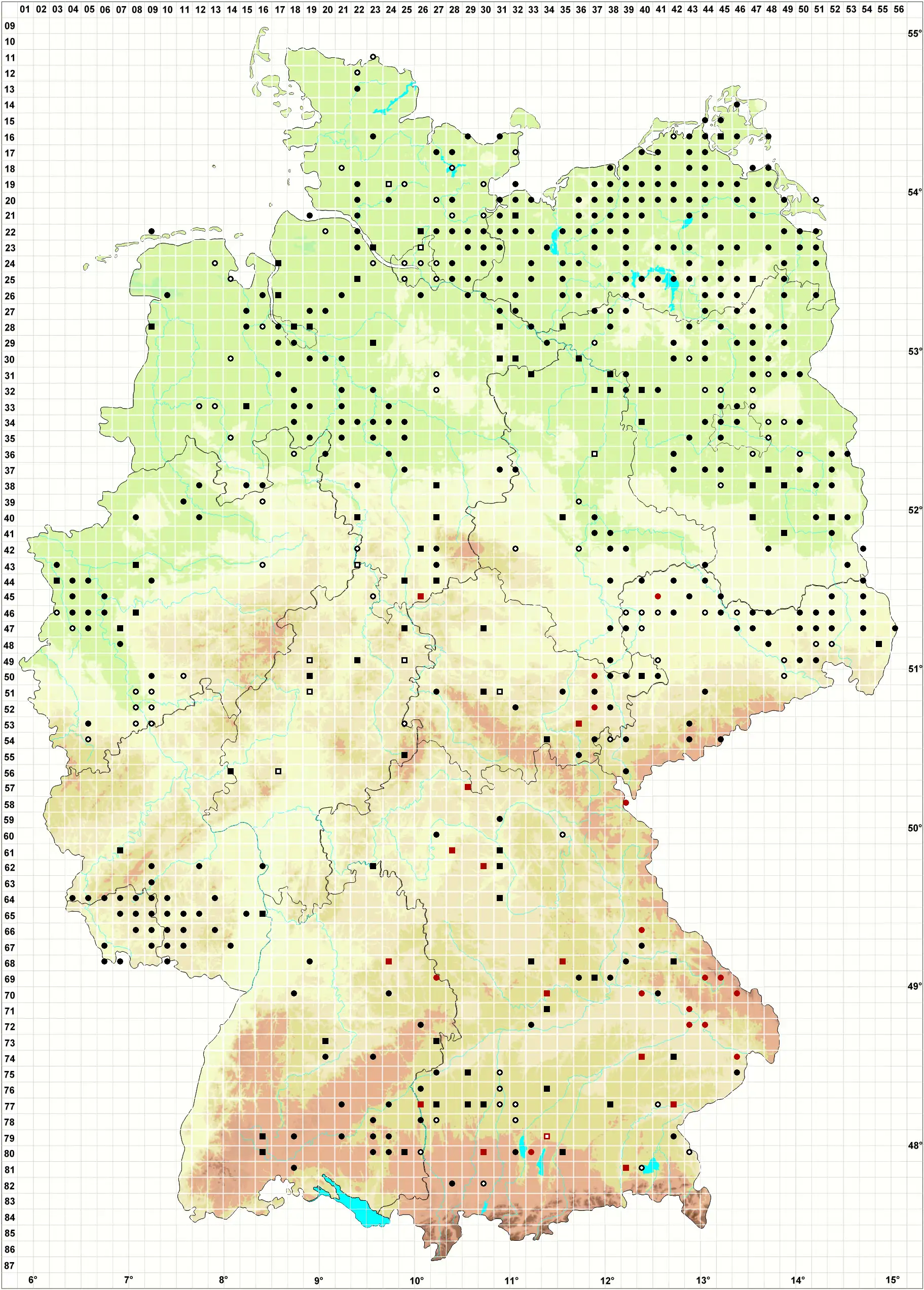
Verbreitung (Meinunger & Schröder 2007) Eine Art des Flach- und Hügellandes, hier durch das ganze Gebiet. In höheren Lagen nur vereinzelt, in den Alpen anscheinend fehlend. Die Karte ist noch in vielen Teilen unvollständig, da die Art oft nicht erkannt oder nicht beachtet wurde. Literaturangaben sind oft unsicher. Wir geben eine Auswahl von uns geprüfter Belege sowie einiger uns zuverlässig erscheinender Literaturangaben: SH: 2128/3 Salzstelle am Traveufer bei Alt Fresenburg, 06.10.1924, F. Koppe (!); 2326/4 Farmsener Tongruben, 6/1909, leg. Wahnschaff als Amblystegium paludosum Hansen, von Loeske als Amblystegium kochii bestimmt (!). Literatur: N. Jensen (1952, die dortigen Angaben zu A. varium var. paludosum dürften hierher gehören); Frahm & Walsemann (1973); Dengler et al. (2000); Siemsen et al. (2000); Siemsen (2001; 2003). MV: Häufig: C. Berg (in litt.). 1645/4, 11.10.1998, WS. NE: 2119/4 Küste N Benfleth, mit Pottia heimii, 28.04.1999, LM; 2417/1 Salzwiese SW Weddemarschen, mit Pottia heimii, 30.04.1999, LM; 2522/4 Auetal S Bargstedt, Nasswiese, 09.05.1999, WS; 2809/1 Bunderneuland, Ackerrand, 12.09.2000, LM; 3032/3 Salzstelle SO Schreyahn, 31.10.1992, WS; Koperski (1998a; 1999; !). STE: 3138/3 feuchte Senke hinter dem Elbdeich in Berge, 23.06.1995, WS; 3237/4 Waldsumpf N Klein Ellingen, 23.06.1995, WS; 4035/4 ehemalige Dorfstelle Luxdorf, mit Drepanocladus aduncus, 03.10.1994, WS; Zschacke (1912); P. Kaiser (1896). BB: Offenbar verbreitet, vergl. Klawitter et al. (2002). 2942/4 Tornowsee, 04.10.1998, WS; 3440/1 Rodewaldsches Luch, 02.04.2001, WS; 3450/2 Mühlteich NW Münchehofe, 08.04.1975, R. Düll (!); 3847/3 Klein Leber-Luch SW Egsdorf, 26.10.1994, WS; 3849/1 Dobra-See, 15.04.2000, WS; 3851/2 Ragow, 10.07.1983, LM; 4047/2 Feuchtstelle im Wald 2 km NO Prierow, 04.06.2000, LM; 4152/1 N Maust, 27.09.1996, LM; J. Klawitter (!); S. Rätzel (!); V. Otte (!); C. Grätz (!). NB: 3815/3 untere Noller Schlucht, Feuchtgrünland, 25.04.1992, M. Koperski (!); 3827/4 Quelle in der Feldmark W Altenhagen, 14.05.2001, F. Jurkutat (!); Koperski (1998); M. Preussing (!); E. Walsemann (!); U. de Bruyn (!). ST: Loeske (1903); Zschacke (1903; 1905); F. Müller (1993). NW: 3812/3 nahe Eltingmühle, 22.03.1993, C. Schmidt (!); 3911/3, April 1997, C. Schmidt (!); 4604/13 Niersaue W Kempen, feuchte Wiese, 14.01.1996, U. Abts (!); 4707/1 R. Düll (!); Solga (1998). HE: 4922/2 Kohlengrube S Ostheim, 01.06.1994, (!!); J. Futschig (FR, !); M. Preussing (!); E. Baier (!); P. Tautz (!). TH: 5040/3 Roter Berg, auf Plattendolomit, 1995, K. Strumpf (!); 5232/3 Humbachsteiche SW Cottendorf, 11.07.1981, LM; 5536/3 Saaldorf, Schlammfläche am Stausee, 05.07.1997 (!!). SN: F. Müller (2004); M. Baumann (!). RP, SL: Lauer (2005); Caspari (2004); J. Werner (!). BW: Nebel in Nebel & Philippi (2001); M. Lüth (!); U. Schwarz (!); 6223/4 Taubertal W Niklashausen, 02.03.1993, (!!); 7327/4 Straßenrand beim Viehhof S von Sachsenhausen, 15.05.2001, LM. BY: 6128/4 Teichrand N Kleingressingen, mit Drepanocladus aduncus und Eurhynchium hians, 30.04.1998, LM; 6431/2 Feuchtwiese O Niederndorf, 17.04.2000, LM; 6839/2 Wiesengräben am Regen O von Nittenau, 01.05.2004 (!!); 6842/2 Waldsumpf N Gadsdorf, mit Amblystegium radicale, 12.06.1996, WS; 6937/3 Schilfsumpf am Mühlbach W Neudeuerling, 21.05.1996, WS; 7134/1 Nasswiese S Schambach, 13.05.1996, LM; 7233/2 Quellhang O von Irgertsheim, 09.05.1996, LM; 7440/4 Feuchte Wiese O von Aham, 20.03.1998, LM; 7442/2 feuchte Wiese N Arnstorf, 16.03.1998, WS; 7634/2 NSG O Weichs, an Salix, 01.04.1998, WS; 7726/2 NSG Illerberg, 17.04.1997, WS; 7727/1 Klosterweiher Roggenberg, 17.04.1997, WS; 7729/2 W von Fischach, 15.1997, WS; 7730/3 Nasswiese S Waldberg, 15.04.1997, WS; 7738/1 Vallei, Erlenbruch, mit A. radicale, 23.03.1998, WS; 7742/1 feuchte Wiese SW Aichberg, 14.03.1998, LM; 7942/1 Bergham, kleiner Seggensumpf, 09.05.1998, WS; 8030/2 Nasswiese NO Waalhaupten, 03.04.1997, LM; 8035/1 Moorgebiet S Dingharting, 18.09.1995, WS; A. Huber (1998); O. Dürhammer (!); R. Lübenau (!).
Bestand und Gefährdung (Meinunger & Schröder 2007) Amblystegium humile wächst meist zwischen anderen Moosen, seltener in geschlossenen Rasen und kann so größere Bestände bilden. Ihre Verbreitung ist noch unzureichend erfasst, besonders steril wird sie leicht übersehen, die auffälligen Sporogone mit langen Seten findet man nur gelegentlich. Die Pflanze findet sich auch an anthropogen geschaffenen oder beeinflussten Standorten. Auf Grund der vorliegenden floristischen Daten und der Standortansprüche scheint die Art im gesamten Untersuchungsgebiet ungefährdet zu sein.

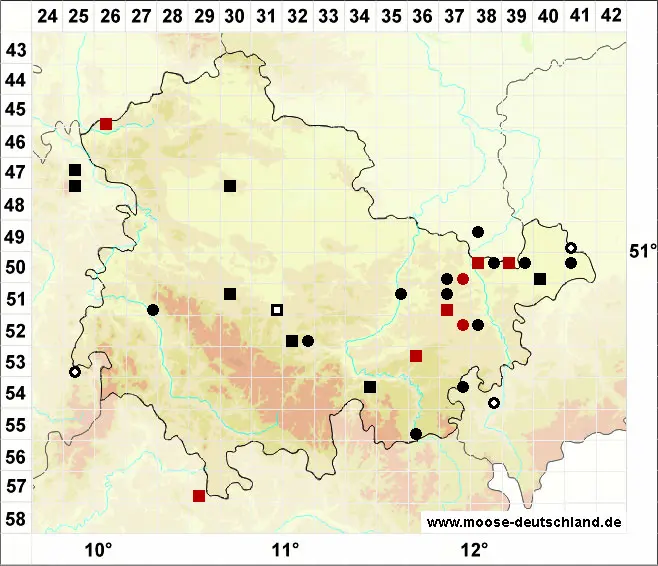
 [ Verbreitung in Deutschland ]
[ Verbreitung in Deutschland ]