Habitat/Ökologie (Meinunger & Schröder 2007) Diese neue Art wird bei Schuster (1980) ausführlich beschrieben. Seine Ausführungen werden von Schumacker & Váña (2000) und Damsholt (2002) übernommen, ohne dass neue Gesichtspunkte hinzukommen. Schuster bezeichnet seine Art selbst als „A rather baffling taxon“ und fasst sie in einem sehr weiten Sinn. Die Art gehört zweifellos in den Cephaloziella arctica-Komplex und ist damit durch Zwischenformen verbunden. Bei unseren Untersuchungen sind uns eine ganze Reihe Proben begegnet, die in diesen Komplex gehören, und wir meinen, die Ansichten von Schuster teilweise einschränken und präzisieren zu können. Das von Schuster als entscheidend herausgestellte Merkmal der scharfen, ein- bis zweizelligen Blattspitze, die mehr oder weniger stark bis hakenförmig nach innen gekrümmt ist, halten wir für völlig ungeeignet. Dieses Merkmal findet sich auch bei Cephaloziella variansalpina, Formen von C. hampeana und anderen Cephaloziella-Arten und kann zur Artabgrenzung nicht verwendet werden. Viel wichtiger sind die sehr stark und auffällig, oft mehrzelligspinulös gezähnten weiblichen Hüllblätter, wie sie in ähnlicher Form in dieser Gattung nur noch bei C. elachista vorkommen. Unterblätter sind an sterilen Sprossen immer deutlich und auffällig entwickelt. Charakteristisch sind ferner die stark spreizenden Perianthmündungszellen. In dieser Merkmalskombination sind die Pflanzen so auffällig, dass jeder, der einmal solche Proben gesehen hat, sie nicht mehr verwechseln kann. Im Habitus entsprechen sie genau der von Schuster gegebenen Beschreibung. Die Pflanzen sind hell- bis blassgrün, teilweise ausgebleicht und C. hampeana ähnlich. Von letzterer Art sind sie sofort durch die überall vorhandenen auffälligen Unterblätter zu unterscheiden. Die meist dünnwandigen bis mäßig verdickten Zellen können gelegentlich stärker verdickt sein, die Pflanzen zeigen bisweilen an den Blatträndern und Sprossspitzen einen roten Farbanflug. Nur Pflanzen mit dieser Merkmalskombination bezeichnen wir als C. uncinata, sie entsprechen weitgehend dem, was Schuster (1980, Fig. 515: 1–9) als „sphagnicola phenotype“ bezeichnet, was etwas irreführend ist. Wir haben Proben sowohl von feuchten, humiden als auch von trockenwarmen Standorten gesehen, die sich immer gleichen, es kann sich also nicht um „Ökoformen“ handeln. Es handelt sich auch keineswegs um eine arktische, sondern um eine zwar offensichtlich seltene, aber auch in Mitteleuropa weit verbreitete Art, die hier bisher nur nicht erkannt wurde. Schon Schuster (1980) weist darauf hin, dass es Mittelformen zu Cephaloziella varians gibt. Das betrifft vor allem Fig. 514, „C. arctica (alpina phenotype)“ und Fig. 515: 10–21, „C. uncinata (uncinata phenotype)“, die sich nach dem derzeitigen Kenntnisstand kaum sicher trennen lassen. Bei diesen Formen sind die weiblichen Hüllblätter nur schwach gezähnt, auch die übrigen Merkmale sind intermediär. Eine von uns gesammelte Probe vom Wendelstein bei Bayrischzell hatte U. Schwarz als C. uncinata bestimmt, was Damsholt bestätigte und unter diesem Namen in Koperski et al. (2000) veröffentlicht wurde. Die Probe hat aber nur eine gekerbte Perianthmündung und schwach gezähnte weibliche Hüllblätter, sie gehört eindeutig zu C. alpina Douin, eine längst bekannte und in den Alpen vermutlich verbreitete Art. Viel schwieriger zu beurteilen sind Formen, die wir auf Kalkschiefer an der Bahn bei Gebersdorf in Thüringen sammelten. Hier wachsen Formen von C. varians und C. uncinata nebeneinander (vergl. Schuster 1980: 160). Man wird künftig nach weiteren trennenden Merkmalen suchen müssen, wir führen vorerst solche Formen unter C. varians. Ökologisch bestehen zwischen Cephaloziella uncinata im hier gebrauchten Sinne und den übrigen Formen von C. varians praktisch keine Unterschiede. Die Pflanzen wachsen auf basenhaltigen, offenen bis halboffenen, mehr oder weniger feuchten Stellen an Wegrändern, in Kiesgruben und Steinbrüchen sowie zwischen anderen Moosen über Felsen.
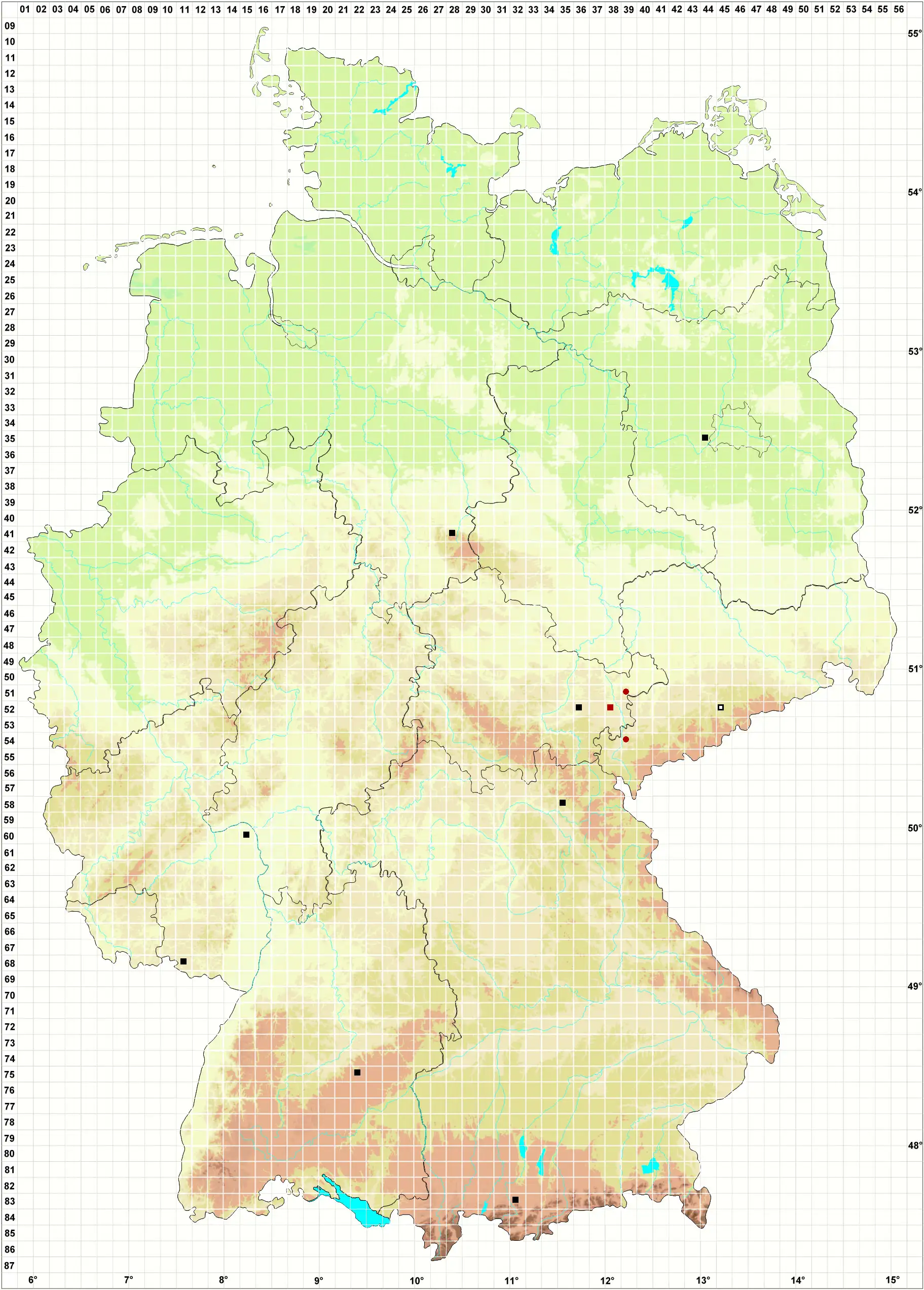
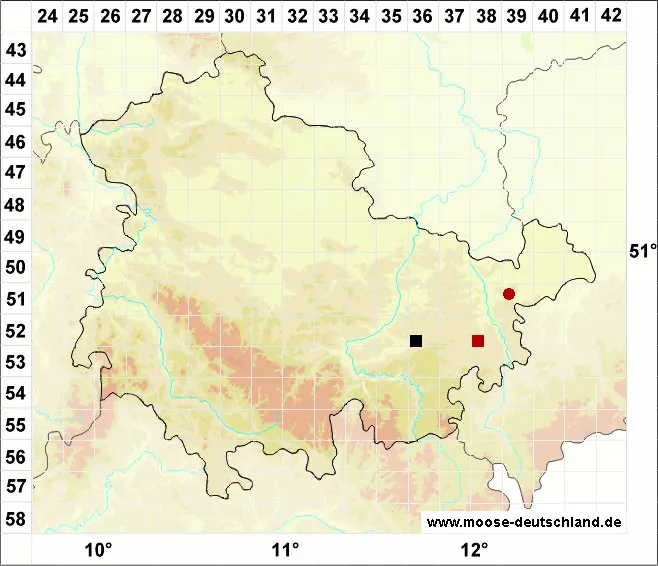
Verbreitung (Meinunger & Schröder 2007) Vereinzelt im Flach- und Hügelland bis in die submontane Stufe. Es werden nur Proben im oben definierten Sinne aufgelistet: BB: 3544/2 NO Fahrland, Abgrabungsstelle S vom Kien-Fenn, basenreicher Sand, mit Encalypta vulgaris, Salvia pratensis und weiteren wärme- und basenliebenden Arten, 11/2001, S. Rätzel & J. Fürstenow (!). NB: 4128/2 Okertal, Rabenklippe, Kalkblock, zwischen Kalkmoosen. 10.09.1999, M. Preussing (!). TH: 5236/3 Drommberg SO Kolba, mit Campylium calcareum, in der Nähe auch Scapania gymnostomophila, 27.09.1970, leg. R. Marstaller, det. LM (JE !); 5238/3 Weidatal 1,5 km SW Loitsch, Felsen, 280 m, 19.08.1981, J. Rettig (!). SN: 5245/3 Lengefeld, auf moosigen Blöcken im schattigen Grunde des Kalkbruches unter den Kalköfen, 640 m, 12.08.1933, leg. W. Flössner (JE !). RP: 6015/1 W Mainz-Lerchenberg im Oberolmer Wald, Waldboden, Mergel, 230 m, 05.06.1995, A. Oesau (!); 6811/333 SW Eppenbrunn, auf einem mit Melaphyr gekiesten Waldweg, an etwas schattiger, leicht feuchter Stelle, 08.08.1993, H. Lauer (!). BW: 7522/1 Ermstal S Bad Urach, beschatteter Trockenrasen, mit Bryum capillare, Frullania tamarisci, Fissidens cristatus, 13.03.1997, M. Preussing (!). BY: 5835/1 Aufgelassener Diabassteinbruch N Stadtsteinach, auf übererdeten, feuchten Steinen, 04.09.2004 (!!); 8332/1 Bayersoien, Ufer des Soiernsees, an leicht schattiger Torfböschung am Rande eines Moorwaldes, mit Campylopus pyriformis, 850 m, 05.07.2000, H. Lauer (!).
 [ Verbreitung in Deutschland ]
[ Verbreitung in Deutschland ]